

TEL:17312606166(魏經(jīng)理)




美鳳力臨床前大動物實驗中心

 17312606166
17312606166
ES細胞打靶技術(shù)
CRISPR/Cas9技術(shù)
在小鼠胚胎干細胞(ES 細胞)中進行 DNA 同源重組,將 ES 細胞重新注射到囊胚腔中,可以形成嵌合胚胎,并能在假孕小鼠體內(nèi)發(fā)育,最終發(fā)育為嵌合體小鼠。這種嵌合小鼠的組織細胞(包括生殖細胞)可以分別來自囊胚的細胞和被注射的ES細胞。因此,通過嵌合小鼠和野生型小鼠的交配,ES 細胞中的遺傳信息就能夠傳遞到后代的小鼠個體中。
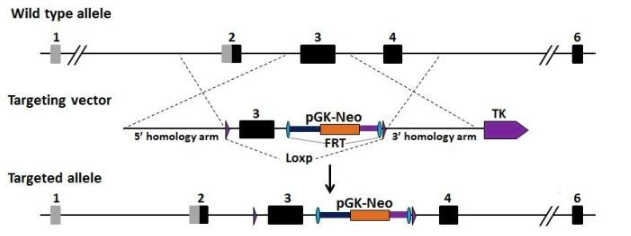
利用ES細胞打靶技術(shù)建立基因修飾小鼠模型的主要技術(shù)路線:
(1)構(gòu)建重組基因載體(正負雙向選擇系統(tǒng))
(2)用電穿孔等方法把重組DNA轉(zhuǎn)入受體ES細胞內(nèi)
(3)用選擇培養(yǎng)基篩選抗藥ES細胞,并利用PCR或或 Southern blot 等方法鑒定發(fā)生正確同源重組的ES細胞
(4)將中靶ES細胞注射到受體囊胚中,將注射后存活的胚胎移植到假孕小鼠子宮中,讓其受孕,待小鼠出生后,通過觀察毛色的嵌合與否和嵌合程度判斷是否獲得嵌合體小鼠。
CRISPR/Cas是細菌和古細菌在長期演化過程中形成的一種高度保守的適應(yīng)性免疫防御系統(tǒng),可用來對抗入侵的病毒及外源DNA。
CRISPR是Clustered regularly interspaced short palindromic repeats(規(guī)律成簇間隔短回文重復)的縮寫。CRISPR簇是一個廣泛存在于細菌和古生菌基因組中的特殊DNA重復序列家族,其序列由一個前導區(qū)(Leader)、多個短而高度保守的重復序列區(qū)(Repeat)和多個間隔區(qū)(Spacer)組成。
Cas則是在CRISPR附近的一個多態(tài)性家族基因,這個家族基因編碼的蛋白均含有可與核酸發(fā)生作用的功能域,并且與CRISPR區(qū)域共同發(fā)揮作用,因此被命名為CRISPR關(guān)聯(lián)基因(CRISPR associated),縮寫為Cas,包括Cas1~Cas10等多種類型。
當細菌抵御噬菌體等外源DNA入侵時,在前導區(qū)的調(diào)控下,CRISPR被轉(zhuǎn)錄成pre-crRNA,然后被加工成一系列短的含有保守重復序列和間隔區(qū)的成熟crRNA,最終識別并結(jié)合到與其互補的外源DNA序列上發(fā)揮剪切作用。
圖3. 細菌CRISPR/Cas系統(tǒng)抵御外來入侵原理示意圖(圖片來自Cell. 2014 Jun 5;157(6):1262-78.)
目前在基因編輯應(yīng)用最為廣泛的CRISPR/Cas9系統(tǒng)是以Cas9蛋白以及向?qū)NA(gRNA)為核心組成。Cas9含有在氨基末端的RuvC和蛋白質(zhì)中部的HNH兩個活性位點,在crRNA成熟和雙鏈DNA剪切中發(fā)揮作用,引起DNA雙鏈斷裂。CRISPR/Cas9的剪切位點位于crRNA互補序列下游鄰近的PAM區(qū)(Protospacer Adjacent Motif)的NGG位點,這種特征的序列在每128bp的隨機DNA序列中就重復出現(xiàn)一次。
圖4. CRISPR/Cas9基本結(jié)構(gòu)原理示意圖(圖片By Mariuswalter - Own work, CC BY-SA 4.
DNA雙鏈斷裂后,在不存在同源模板的情況下發(fā)生非同源性末端接合(Non-homologous end joining,NHEJ)修復斷裂DNA。通常在Cas9剪切位點附近造成小片段堿基的插入或缺失。如果隨機修復發(fā)生在基因的編碼外顯子中,則往往會導致讀碼框發(fā)生移碼突變,最終使靶基因無法正常轉(zhuǎn)錄翻譯。當DNA斷裂后,細胞核內(nèi)同時存在與損傷DNA同源的DNA片段,那么則可通過同源介導的雙鏈DNA修復(Homology directed repair ,HDR)在目的位點引入外源DNA片段,從而達到片段敲入或編輯的效果。
利用CRISPR/Cas9系統(tǒng)的這些特性,CRISPR/Cas9途徑獲得基因修飾小鼠模型的主要技術(shù)路線:
(1)我們可以針對基因組特定序列設(shè)計向?qū)NA(gRNA)
(2)體外轉(zhuǎn)錄gRNA以及編碼Cas9蛋白的mRNA
(3)將gRNA、Cas9 mRNA注射到小鼠受精卵中
(4)注射后的受精卵移植到假孕母鼠輸卵管中,受精卵發(fā)育成F0代小鼠。
由于高等真核細胞內(nèi)外源DNA與靶細胞DNA序列自然發(fā)生同源重組的機率非常低,約為百萬分之一,要把基因敲除成功的細胞篩選出來是一件非常困難的工作。因此,需要借助一些篩選標記來解決這一關(guān)鍵問題。
“+” 正篩選標記:通常是藥物抗性基因,最常用的就是Neo(新霉素抗性基因)。通過構(gòu)建打靶載體,將帶有目的片段的外源DNA序列連同正篩選基因 neo 通過 DNA 序列間的同源重組替代基因組中需要敲除或修飾的片段,達到敲除或敲入特定基因的目的。正篩選基因 neo的表達賦予重組 ES 細胞具有抵抗細胞毒性藥物 G418的特性,使細胞在藥物篩選下存活并形成細胞克隆。
“-” 負篩選標記:為減少因隨機插入產(chǎn)生的藥物抗性克隆對篩選的干擾,可以在打靶載體同源重組區(qū)域的外側(cè)加上負篩選基因,通常是細胞毒性相關(guān)基因。例如來自皰疹病毒的胸腺嘧啶激酶(TK)基因,該基因可以將對細胞無毒的核苷類似物 Ganc 進行磷酸化,從而產(chǎn)生對細胞有毒的核苷酸類似物。 當打靶載體在細胞內(nèi)進行同源重組時,TK 基因由于在同源重組區(qū)域的外側(cè),因此不會整合到細胞染色體上,隨著細胞的生長和分裂 TK 基因發(fā)生自然降解和稀釋,從細胞中消失。因此這類細胞不會因為 Ganc 的存在而死。但如果發(fā)生打靶載體隨機插入整合到 ES 細胞染色體上的情況,那么TK 基因很可能也會同時整合上去,結(jié)果導致這類非正確同源重組的ES細胞在 Ganc 的存在下生成有毒的核苷酸類似物而致死。
| 上一篇:小鼠胚胎首次在太空中生長 | 下一篇:經(jīng)典的模式生物都有哪些?大腸桿菌、擬南芥、近交系小鼠、果蠅 |
|
|
|
|
